COMMENT: Mr Armstrong, THANK YOU VERY MUCH !!! for telling your readers of the true nature of the Hamas; God bless you for telling the truth. Millions of the impoverished people of Gaza live in fear of these thugs. History shows Jews, Arabs, and Christians CAN, yes, CAN live together in peace, and it is the politicians or extremist groups, inhuman groups like Hamas, that kindle the fire of hatred.
Human beings can be both strong and weak at the same time. When they fall into the trap of hatred, it is very difficult to get out of it; see what happened in Northern Ireland for many years. No one in Israel gets up in the morning wishing to ‘take care’ of the Palestinians; they can take care of themselves if only their leaders will not rob them blind and stuff their brains with poisonous ideology.
George Soros: This creature that looks to have a human shape is not a human being at all; it is one of the most despicable creatures on the planet Earth. God delays taking him back, I guess, because even God will be annoyed to have him around up there!
Wishing you always the best, Ronnie
REPLY: I have been throughout the region. I have driven through the West Bank and spoken to both Israelis and Palestinians. I have explored the entire area and even took the first flight between Israel and Egypt when relations began. The common people get along. This documentary, The Heart of Jenin, is a 2008 documentary film written and directed by Marcus Vetter and Leon Geller. This film is important to watch for it reveals the story of Ismael Khatib from Jenin, a Palestinian whose son was shot by Israeli soldiers by mistake because he had a toy gun.
Instead of seeking revenge, he donated his son’s organs to Israeli children. In April 2010, The Heart of Jenin won the German Film Award for Best Documentary Film. What this film shows is the common people – not the political hatred of extremists who live for violence no different than our American Neocons who love war and have no problem sacrificing your children for their political agendas. When Field Marshall Goring was put on trial at Nuremberg, he explained something that is so true, but it is never discussed in school and certainly never by mainstream press, which has become the cheerleader for World War III.
Goring was absolutely correct. Those in charge love war and manipulate the people for their own objectives. Hamas wants Israel to kill Palestinian civilians for their own agenda. Suffice it to say I also have a source who knew those directing young Arabs to do suicide bombings. As the source told me, those people would NEVER kill themselves – they always preach others to do that for their agenda.
Our CI wanted to kill American civilians so they could blame it on Cuba to justify invading the island. President Kennedy said no way. That is why the CIA refuses to release all the Kennedy documents because most people believe they took out Kennedy.
THE MURDER OF JFK
I believe the entire Watergate Affair was set up by the CIA because Nixon knew the CIA had assassinated Kennedy. The Watergate scandal involved the five men caught on June 17, 1972, burglarizing the Democratic National Committee’s headquarters in the Watergate complex. Their two handlers were E. Howard Hunt of the CIA and G. Gordon Liddy of the FBI. They were Nixon campaign aides. Nixon wanted to end Vietnam and he knew the CIA was behind the Kennedy Assassination because they wanted war and BELIEVED that Russia was involved.
Before the Neocon Robert Macnamara died, he came out and admitted that they were wrong about Vietnam. That was nice, but 58,000 Americans died for that mistake, and untold millions of Vietnamese.
We are always looked down upon as their pawns to be played. If it is Hamas, CIA, or any government body, they will ALWAYS act in their own self-interest, which is NEVER for the benefit of the people. The only solution to war is to throw the warmongers in a closed room and let them fight it out and leave the common people alone.
I explained that Romania is becoming a World War III training battleground for NATO. There are currently 40,000 troops on the ground throughout Eastern Europe, with more set to arrive. Colonel Vincent Minguet, the commander of the NATO Combat Group in Romania, believes the training will last up to FIVE years. Stronger militaries are training their allies now in preparation for what is to come.
This is expanding beyond Eastern Europe. France announced that they are welcoming 2,000 Ukrainian troops. French President Emmanuel Macron was hesitant at first, and only permitted 40 Ukrainian soldiers to be trained on French soil since February. As Zelensky and those holding his strings beg for more aid and a full-on escalation, world leaders are folding to his demands.
Under Operation Interflex, the UK has coached 10,000 Ukrainian fighters. The program puts soldiers through an intense five-week training program to learn basic skills before being returned to the frontline.
Ireland has been tasked with helping Ukraine learn to dismantle explosives and IEDs. The US has training facilities for Ukrainian soldiers in Germany, while the UK has opened up air bases for Canadian forces. Every nation is preparing for endless war. “We are probably, unfortunately, in the long haul when it comes to the war against Russia to stop Putin’s aggression against Ukraine. Therefore, we need also long-term planning and training,” Danish Foreign Minister Jeppe Kofod admitted.
there was a sabotage act on the German railroad company „Deutsche Bahn“. Two cables in different locations were cut on Saturday morning (October 8th, 2022). Result: No trains were moving in all of northern Germany. This must have been accomplished by someone highly sophisticated and with insider knowledge. First, someone blew up Nordstream 1 and 2 pipelines, now someone sabotages Deutsche Bahn. Are we (Germany) already in a war?
Kind regards from Germany,
Michael
REPLY: There is a very strong rising element of this Climate Change Zealots. They are out to destroy all fossil fuel production. This element is very strong in Europe, although it is rising in the United States as well. They have no idea what they are unleashing upon the world. Yet, there is no talking to these people. They have been totally brainwashed. These people are totally insane and they see humanity as the problem.
Die großflächigen Ausfälle im Zugverkehr in Norddeutschland gehen nach Aussage der Bahn auf Sabotage zurück. »Aufgrund von Sabotage an Kabeln, die für den Zugverkehr unverzichtbar sind, musste die Deutsche Bahn<https://www.spiegel.de/thema/deutsche_bahn/> den Zugverkehr im Norden heute Vormittag für knapp drei Stunden einstellen«, sagte eine Sprecherin der Bahn dem SPIEGEL. Die zuständigen Sicherheitsbehörden hätten die Ermittlungen aufgenommen.
Nach SPIEGEL-Informationen fiel das Funk-Kommunikationsnetz der Bahn am Samstagmorgen gegen 6.40 Uhr komplett aus. In Sicherheitskreisen hieß es, zuvor hätten Unbekannte bei Berlin-Karow eine für das Kommunikationsnetz entscheidende Datenleitung in einem Kabelschacht durchtrennt. Auch bei Dortmund wurde kurz vor dem Totalausfall ein ähnlicher Sabotageakt in einem Kabelschacht festgestellt, so ein Ermittler. Ob die beiden Taten in Zusammenhang stehen, werde derzeit noch geprüft.
Für eine gezielte Sabotage-Aktion, so an den Ermittlungen beteiligte Beamte, brauche man Informationen über das Netzwerk der Bahn und wie man es lahmlegen kann.
Empfohlener externer Inhalt
An dieser Stelle finden Sie einen externen Inhalt von Twitter, der den Artikel ergänzt und von der Redaktion empfohlen wird. Sie können ihn sich mit einem Klick anzeigen lassen und wieder ausblenden.
Ich bin damit einverstanden, dass mir externe Inhalte angezeigt werden. Damit können personenbezogene Daten an Drittplattformen übermittelt werden. Mehr dazu in unserer Datenschutzerklärung. <https://www.spiegel.de/datenschutz-spiegel>
Im Lauf des Vormittags wurde die heftige Störung laut Bahn behoben. Allerdings seien noch Beeinträchtigungen im Laufe des Tages möglich.
Mehrere Quellen aus Bahn-Kreisen bestätigten dem SPIEGEL am Vormittag, dass der Grund der Bahnstörung ein flächendeckender Ausfall des von der Bahn genutzten Funknetzes GSM-R in Norddeutschland gewesen sei. Über dieses Netz läuft nicht nur der Sprechfunk zwischen Leitstellen und Zügen, sondern auch die digitale Übermittlung von Fahrplandaten. Die Kommunikation über GSM-R sei eine »zentrale Schnittstelle zwischen Fahrzeugen und Infrastruktur«, hieß es. Ein Kontakt zu Lokführern und Personal war zwischenzeitlich nicht möglich.
Empfohlener externer Inhalt
An dieser Stelle finden Sie einen externen Inhalt von Twitter, der den Artikel ergänzt und von der Redaktion empfohlen wird. Sie können ihn sich mit einem Klick anzeigen lassen und wieder ausblenden.
Ich bin damit einverstanden, dass mir externe Inhalte angezeigt werden. Damit können personenbezogene Daten an Drittplattformen übermittelt werden. Mehr dazu in unserer Datenschutzerklärung. <https://www.spiegel.de/datenschutz-spiegel>
Den Angaben zufolge war nicht nur der Fernverkehr, sondern auch der Regionalverkehr von der Störung betroffen.
Auch im internationalen Fernverkehr fielen Züge aus: Auf der Strecke zwischen Berlin und Amsterdam entfielen IC-Züge komplett. IC-Züge von und nach Kopenhagen und Aarhus endeten in Padborg.
Die Bahn hob als Folge der Störungen die Zugbindung bereits gekaufter Fernverkehrstickets auf. Wer eine Reise für Samstag gebucht habe, könne diese »bis einschließlich sieben Tage nach Störungsende flexibel nutzen. Sitzplatzreservierungen können kostenfrei storniert werden«, erklärte das Unternehmen <https://www.bahn.de/service/fahrplaene/aktuell> . Wer nicht mehr reisen möchte, kann sich den Ticketpreis erstatten lassen.
Der Ausfall traf nicht nur Wochenendpendler, sondern auch Familien auf dem Weg in den Urlaub: In Hamburg<https://www.spiegel.de/thema/hamburg/> und Schleswig-Holstein haben am Samstag die Herbstferien begonnen.
Anmerkung: In einer früheren Version war von dem Funknetz GSR-M die Rede – tatsächlich heißt das von der Bahn genutzte Netz aber GSM-R. Wir haben die entsprechende Stelle korrigiert.
Alison Morrow Published originally on Rumble on October 7, 2022
#FDA #Lawsuit #Doctors A group of doctors is suing the FDA, HHS and the men in charge of each agency for interfering in their ability to treat patients. They say the federal departments lied about Ivermectin.
Canada is finally ending the absurd COVID vaccine mandates. No, Trudeau did not come to his senses. The truth of the matter is that the majority of people dying from COVID are the triple and quadruple vaccinated. In fact, they account for 90% of COVID-related deaths in the past three months.
There is true mass psychosis surrounding the vaccination. It does not prevent transmission or infection. Despite that being a known fact, politicians are eager to promote the vaccine, along with the media that continues to push boosters on every platform available. People are bringing their babies and elderly loved ones to get the latest vaccine with lines out the door. What happens now when the mask falls and people realize the truth? There are people crying online that it is “too soon” to remove the vaccine requirement. It is cognitive dissonance. They cannot allow their brains to process that they were lied to for years. The vaccine only weakened people’s immune systems and made them more susceptible to COVID.
IT WAS NEVER SAFE AND EFFECTIVE.
I congratulate the Canadian Truck Convoy and all others who risked their liberty and lives to protest this false narrative. The silent majority has won, but at what cost? Countless lives were destroyed over this mandate. People lost their livelihoods and homes. Some who I personally know were forced to flee Canada as they were not willing to risk their health. Businesses were ruined. Families were separated as the demonized unvaccinated could not even enter Canada. Lives were completely uprooted due to this vaccine mandate imposed by the Trudeau Administration.
And now they want to pretend everything is ok and sweep it all under the rug. No. We should remember what Trudeau and the globalists did to the people. The unvaccinated were gas lighted and banished from society for years. The people were threatened in every way possible until they agreed to submit to the government’s order. We now see how much control the government has over the people. Public confidence will never be the same.
Geert Vanden Bossche, DVM, PhD General Manager at Voice for Science and Solidarity | The biggest challenge in vaccinology: Countering immune evasion posted originally on TS New on Aug. 1, 2022
Exposure of a highly C-19 vaccinated population to monkeypox virus (MPV) spilling over from an animal reservoir promotes asymptomatic human-to-human transmission in susceptible sexual minority communities (SMCs). MPV infection in SMCs could therefore evolve more infectious viral variants that spread to all parts of a highly C-19 vaccinated population and thereby prevent establishment of herd immunity
Increasing numbers of outbreaks of human monkeypox have been reported from across central and west Africa over the last 3-4 decades. Zoonotic infection with MPV in the current setting of limited smallpox vaccination and little orthopoxvirus immunity[1] in several parts of the world renders human populations more susceptible to contracting monkeypox disease. MPV has therefore been considered a well-suited candidate for a global epidemic.
As productive poxvirus infection is mostly symptomatic and viral transmission almost exclusively occurs through close contact with an infected animal or person or via virus-contaminated objects, such as bedding or clothing, it has been generally acknowledged that naturalepidemic outbreaks in humans can largely be contained through basic infection-prevention measures (including good hygiene practices). Unless the viral infection rate is high (e.g., in densely populated areas and poor [environmental] hygiene conditions), it is difficult to imagine how MPV could evolve to adapt to the human population, let alone how it could ignite a multi-country epidemic or even a pandemic in countries where MPV is not an endemic zoonosis. Pandemics typically occur with pathogens that cause so-called acute self-limiting infection, meaning that they have the potential to spread asymptomatically before inducing a type of natural immunity that prevents productive infection upon subsequent exposure and, therefore, generates herd immunity. Whereas until recently many still tended to believe that the threat of a globally spread MPV was a myth, cases are now being reported globally (at least in all highly C-19 vaccinated parts of the world) to the extent that WHO has now declared MPV a health emergency of international concern—all of this has happened within just a few months. This does not provide enough time for population-level innate immunity to become sufficiently trained to turn MPV infection, which is typically symptomatic (so-called ‘acute self-limiting viral disease’, ASLVD) into an infection that is predominantly asymptomatic (so-called ‘acute self-limiting viral infection’, ASLVI) and can therefore much more easily spread between people. On the other hand, adaptation of a virus to a new host population never implies natural selection of less infectious viral variants, on the contrary. If neither viral evolution nor immune training is responsible for shifting symptomatic into asymptomatic viral transmission (thereby allowing MPV to spread more efficiently from person-to-person and eventually become a pandemic), other non-evolutionary disease-mitigating influences must be considered. As spread of MPV is now particularly expanding in countries with high C-19 vaccine coverage rate and as ASLVD-enabling viruses that are predominantly transmitted through close contact do not spread rapidly, there must be a link between the type of population-level immunity in highly C-19 vaccinated populations and the rapid expansion in prevalence of MPV cases. It’s also important to note that—so far— MPV disease symptoms in these populations have been rather ‘mild’ and predominantly manifest in individuals from the gay and bisexual male community. This already suggests that sexual contact, especially when the latter is at risk of traumatizing the skin or mucosa (e.g., in case of anogenital intercourses), facilitates symptomatic MPV infection.
While I cannot unambiguously prove this, I strongly believe that the sudden emergence of a significant number of (mild) cases of MPV in highly C-19 vaccinated countries is not purely coincidental but related to enhanced activation of broadly reactive, MHC-unrestricted CD8+ T cells in vaccinees. I have previously reported on how a universal CTL (cytotoxic T lymphocyte) epitope can facilitate elimination of host cells infected with ASLVI- or ASLVD-enabling glycosylated viruses and thereby allow recovery from disease, however without inducing immunologic memory (https://www.voiceforscienceandsolidarity.org/scientific-blog/epidemiologic-ramifications-and-global-health-consequences-of-the-c-19-mass-vaccination-experiment). More specifically, MHC-unrestricted CD8+ T cells that now increasingly prevent C-19 disease in healthy vaccinees are the same as those required to abrogate productive infection with other glycosylated viruses that have evolved reduced susceptibility to our innate immune system[2], including poxviruses (https://www.frontiersin.org/articles/10.3389/fimmu.2021.740223/full).
Given the enhanced immune activation of pathogen-nonspecific CTLs[3] in C-19 vaccinees, MPV infection in C-19 vaccinees is likely to become abrogated at an early stage of productive infection, thereby dampening productive MPV infection and potentially causing asymptomatic/ mild infection in sexual minority communities (SMCs) of a highly C-19 vaccinated population. Consequently, MPV infection may even fail to induce MPV-neutralizing antibodies (Abs) in vulnerable[4], C-19-vaccinated individuals that are immunologically naïve to MPV (i.e., today persons younger than 45 to 55 years of age, depending on the country). However, it is reasonable to assume that asymptomatic MPV infection may elicit short-lived, low affinity anti-MPV Abs in these individuals (as has, for example, been reported in case of asymptomatic infection with SARS-CoV-2 (SC-2; https://www.medrxiv.org/content/10.1101/2020.06.22.20137141v2.full.pdf). As asymptomatic infections promote viral transmission within these minority communities, the infection rate of MPV in this vulnerable subpopulation is likely to grow over time. This rise in viral infection rate will subsequently increase the likelihood for previously asymptomatically infected persons from SMCs to become re-infected while titers of their short-lived, low-affinity anti-MPV antibodies are still relatively high. Binding of such low-affinity, non-neutralizing Abs to the virus is thought to enhance viral infectiousness and could thereby cause a disease outbreak in these communities. It is, therefore, reasonable to expect that the proportion of vulnerable individuals who develop virus-neutralizing Abs (i.e., upon their recovery from MPV disease[5]) in the C-19 vaccinated part of the population will increase over time. However, in vulnerable, non-C-19-vaccinated individuals, trained innate immune cells (i.e., NK cells) are likely to prevent MPV from breaking through this first line of immune defense and would therefore largely prevent priming of virus-neutralizing Abs. For the time being, symptomatic manifestations in highly C-19 vaccinated populations are predominantly mild and mostly occurring in SMCs. This suggests that even in cases of symptomatic infection, viral clearance via innate or adaptive cytolytic immune cells (in the case of non-C-19 vaccinated or C-19 vaccinated, respectively) is still effective enough to prevent more problematic symptomatology in most cases.
Rising virus-neutralizing Ab titers can only prevent monkey disease but not viral infection. Hence, re-exposure to MPV of C-19 vaccinated individuals who are in the process of seroconverting promotes natural selection of more infectious MPV immune escape variants while fostering asymptomatic transmission and thereby contributing to a further rise in viral infectious pressure. Due to the steadily growing infection rate in SMCs of highly C-19 vaccinated populations, the overall MPV-neutralizing Ab response in these communities is likely to exert suboptimal immune pressure on viral infectiousness and can therefore be expected to drive dominant circulation of naturally selected, more infectious MPV immune escape variants. Based on all the above, the enhanced infection rate mediated by asymptomatic transmission of MPV in SMCs of highly C-19 vaccinated populations is likely to increase the probability of adaptive evolution of MPV in these communities. It is, therefore, critical to monitor the selective landscape of MPV as unfolded in SMCs of highly C-19 vaccinated populations in order to verify whether the evolutionary trajectory is shifting towards promoting natural selection and expansion of immune escape variants that are more infectious (as is smallpox virus, for example).
Vaccination of vulnerable groups (SMCs) against MPV is likely to accelerate adaptive evolution of MPV in highly C-19 vaccinated populations and could thereby raise the incidence of (severe) MPV disease in vulnerable subsets of non-C-19-vaccinated individuals and ignite multi-country epidemics of MPV in non-C-19-vaccinated animal and human populations that are immunologically naïve to orthopoxvirus
Several countries are now about to start vaccination campaigns targeted at people who are at risk of contracting monkeypox disease using live attenuated, replication-incompetent smallpox vaccine. Both, individuals from SMCs engaging in high-risk sexual behaviors for MPV infection and close contacts of monkeypox cases (including very young children, pregnant women, elderly or immunocompromised individuals) are eligible for MPV vaccination (https://www.ecdc.europa.eu/sites/default/files/documents/Monkeypox-multi-country-outbreak.pdf). Live attenuated, replication-incompetent orthopox (e.g., smallpox) vaccines prime virus-neutralizing Abs in the vast majority of both vaccinated and non-vaccinated individuals (i.e., individuals < 50y). However, unlike live attenuated replication-competent orthopox vaccines[6], they do not train cell-based innate immunity. There can be no doubt that vaccination in the context of more infectious circulating MPV variants will further promote natural selection and dominant propagation of even more infectious immune escape variants and thereby allow MPV to evolve into a human pathogen exhibiting an even higher level of infectiousness (comparable to smallpox?). This situation is reminiscent of that which has been responsible for driving adaptative evolution of more infectious SC-2 (SARS-CoV-2) variants following C-19 mass vaccination campaigns. The evolutionary dynamics of MPV will only be expedited when vaccine coverage rates grow; they could eventually modify the current mode[7] and course of chain of MPV transmission such as to asymptomatically spread to all parts of a homogenously mixed, highly C-19 vaccinated population. This would increase the risk for C-19 unvaccinated subjects to contract MPV disease, especially for those who are particularly vulnerable to MPV disease because of Ab-mediated enhancement of viral infectiousness or enhanced susceptibility to MPV infection due to risky (sexual) behavior (see further below). Because of asymptomatic transmission, highly C-19 vaccinated populations would serve as a human reservoir of more infectious MPV immune escape variants.
Spill-over of more infectious MPV variants to populations that are immunologically naïve to orthopoxvirus is likely to trigger epidemics of MPV in poorly C-19 vaccinated countries
It is reasonable to assume that populations which do not ‘benefit’ from hyperactivation of cytotoxic CD8+ T cells will become more susceptible to productive infection with new, more infectious MPV variants (https://www.voiceforscienceandsolidarity.org/scientific-blog/epidemiologic-ramifications-and-global-health-consequences-of-the-c-19-mass-vaccination-experiment). This applies, for example, to several different animal populations as well as to human populations in poorly C-19 vaccinated countries (e.g., in Africa). Asymptomatic infections in highly vaccinated C-19 countries are likely to promote spill-over events involving transmission of more infectious MPV variants from these highly C-19 vaccinated human reservoirs to vertebrate animals (possibly even including livestock) and poorly C-19 vaccinated human populations that are immunologically naïve to orthopoxvirus. Asymptomatic transmission of more infectious MPV variants can also become problematic for the C-19 unvaccinated in highly C-19 vaccinated countries, particularly for C-19 unvaccinated children and vulnerable people (e.g., part of SMCs) who are immunologically naïve to orthopoxvirus.
In young children, rapid re-infection subsequent to previous asymptomatic MPV infection by more infectious MPV variants is likely to entail a rise in cases of Ab-dependent enhancement of MP disease[8] whereas risky sexual behavior renders individuals from SMCs more susceptible to viral infection. One can therefore expect the incidence rate of monkeypox disease to increase in both, non-C19-vaccinated children and SMC members.
Previous vaccination with smallpox (i.e., cowpox) vaccines will likely improve protection from MPV disease in the non-C-19-vaccinated but not in the C-19 vaccinated.
While recall of Abs induced by vaccination against smallpox virus in the past will provide an additional layer of natural immunity in the unvaccinated, repetitive recall of Spike (S)-specific infection-enhancing Abs[9] in C-19 vaccinated individuals by circulating SC-2 variants will allow the latter to outcompete other glycosylated pathogens for internalization into mucosa-resident dendritic cells, thereby reducing or potentially even preventing recall of previously smallpox vaccine-induced Abs. This would imply that older (> 45-50y) C-19 unvaccinated individuals are likely to benefit from their smallpox-vaccination in the past whereas their C-19 vaccinated peers may not. However, as already mentioned, the infection can be expected to be largely asymptomatic/ mild in the vast majority[10] of vaccinated and unvaccinated individuals in highly C-19 vaccinated populations, even in the absence of previous smallpox vaccination.
No child should be vaccinated against monkeypox during this C-19 pandemic
But even replication-competent smallpox vaccines can put the child’s health at risk. Akin to all other live attenuated & replication-competent vaccines (e.g., childhood vaccines), these vaccines are known to come with a risk of side-effects:
“Health complications can occur after receiving the vaccine, and the risk of experiencing serious side effects must be weighed against the risk of experiencing a potentially fatal smallpox infection. The vaccine may cause myocarditis and pericarditis, which are inflammation and swelling of the heart and surrounding tissues and can be very serious. Based on clinical studies, myocarditis and/or pericarditis occur in 1 in 175 adults who get the vaccine for the first time” (https://www.fda.gov/vaccines-blood-biologics/vaccines/acam2000-smallpox-vaccine-questions-and-answers).
Stated bluntly, vaccination of young children against MPV is at risk of provoking life-threatening disease.
Overall conclusion
The vast majority of C-19 vaccinees and C-19 unvaccinated individuals in highly C-19 vaccinated populations develop asymptomatic (or very mild) infection upon exposure to MPV. However, close and disruptive physical contact may promote viral entry through broken skin/ mucosa and is therefore more likely to cause symptomatic infection. Whereas strong training of cell-based innate immunity is likely to prevent productive infection of C-19 unvaccinated persons in highly C-19 vaccinated populations and contributes to herd immunity, hyperactivated cytolytic CD8+ T cells in C-19 vaccinated individuals can only enhance abrogation of productive infection, resulting in substantial mitigation of disease symptoms.
Due to the current advanced stage of the evolutionary trajectory of the C-19 pandemic in highly C-19 vaccinated SMCs, MPV is likely to evolve more infectious/ pathogenic variants. Public health authorities in several highly C-19 vaccinated countries have now started rolling out MPV vaccination campaigns targeted at SMCs. MPV vaccination in the ‘at risk’ groups typically use live attenuated, non-replicating smallpox vaccines. Although these vaccines are much less problematic in terms of vaccine-induced side effects (they have even been approved for use in immunocompromised or immunodeficient people), they can only prevent orthopox (including smallpox) disease—not productive infection. As the type of protection conferred by these vaccines is solely based on the induction of antigen-specific, virus-neutralizing Abs, MPV vaccination programs using this type of vaccines will inevitably expedite adaptive evolution of MPV and hence, further promote dominant circulation of more infectious immune escape variants. Consequently, even small-scale deployment of live attenuated, non-replicating orthopox vaccines targeted at preventing disease in vulnerable individuals are highly problematic in that they have the potential to rapidly turn highly C-19 vaccinated populations into a human reservoir for asymptomatic transmission of more infectious MPV variants to poorly C-19 vaccinated populations that are immunologically naïve to orthopoxvirus. Viral transmission from these reservoirs is therefore at risk of igniting multi-country epidemics in poorly C-19 vaccinated countries while increasing the risk of Ab-dependent enhancement of disease in young C-19 unvaccinated children and individuals at high risk of exposure to MPV (due to risky behavior) living in highly C-19 vaccinated countries.
Given the current epidemiologic situation, mandatory vaccination against monkeypox cannot be justified, regardless of C-19 vaccination status. In C-19 vaccinated populations, current vaccination campaigns will only promote further expansion of more infectious MPV variants. But even in non-C19-vaccinated countries, vaccination is not a reasonable option. This is because poxvirus epidemics do not generate herd immunity sensu stricto[12] and prevention, therefore, of world-wide poxvirus epidemics is only possible when the virus can be eradicated. However, eradication is only feasible provided there are no asymptomatic reservoirs and a global mass vaccination program is conducted with vaccines that are capable of preventing productive infection. The first condition is obviously not fulfilled since highly vaccinated countries now serve as asymptomatic reservoirs for MPV. The second condition cannot be fulfilled either since this would require usage of replication-competent vaccines, ideally in a pre-exposure prophylactic setting (or at least within a few days after suspected exposure). However, even replication-competent smallpox vaccines would not enable protection from productive infection by more infectious MPV immune escape variants for the latter will not be a good match for the vaccinal Abs and could, therefore, expedite propagation of more infectious variants in non-C-19 vaccinated populations too. Furthermore, side-effects caused by the existing replication-competent smallpox vaccines may raise additional concerns in regard of vaccine safety.
In conclusion, no C-19 unvaccinated person should engage in sexual behavior that is at risk of enhancing MPV infectiousness (e.g., anogenital intercourses) or be vaccinated with zoonotic orthopoxvirus types once human-to-human transmission of antigenically shifted (i.e., more infectious) MPV variants is occurring!
The current MPV pandemic is to be considered an indirect consequence of the unfortunate C-19 mass vaccination program and does not yet constitute a public health emergency of international concern. However, each vaccination program that uses non-replicating vaccines targeted at immunologically naïve ‘at risk’ communities to fight ASLVI-enabling glycosylated viruses[13] will expedite the expansion in prevalence of more infectious immune escape viral variants. This is why the MPV vaccination campaigns that are currently kicked off are not only likely to have a detrimental impact on individual health (particularly in C-19 unvaccinated children and vulnerable people) but should also be considered at risk of provoking a true public health emergency of international concern.
However, as far as highly C-19 vaccinated countries are concerned, the evolution of MPV towards establishing an asymptomatic reservoir of more infectious MPV variants is merely a ‘side-effect’ of the ongoing evolutionary trajectory of SC-2 in these countries. I therefore predict that the imminent detrimental health consequences of the C-19 mass vaccination program will soon obviate the need for further speculation on how the MPV pandemic/ multi-country epidemic is going to evolve in industrialized countries and, therefore, in third-world countries.
POSTSCRIPTUM
Vaccination of vulnerable groups against zoonotic influenza virus (MPV) in a highly C-19 vaccinated population will drive adaptive evolution of zoonotic influenza virus and ignite multi-country epidemics in C-19 unvaccinated countries
The immunological mechanisms underlying asymptomatic transmission of MPV from highly C-19 vaccinated populations to immunologically orthopoxvirus-naïve, C-19 unvaccinated individuals or poorly C-19 unvaccinated populations also largely apply to a zoonotic influenza virus. This is to say that vaccination (with a non-replicating zoonotic flu vaccine) of a C-19 vaccinated subpopulation that is at high risk of contracting zoonotic influenza infection is prone to further promoting the expansion of zoonotic flu virus and causing (severe) influenza disease in vulnerable people from the C-19 unvaccinated part of the population.
Which individuals are to be considered vulnerable to zoonotic influenza virus?
Whereas orthopoxviruses originating from various animal species induce cross-neutralizing Abs, influenza viruses from animal species do not induce broadly cross-neutralizing Abs. Individuals who received smallpox (i.e., cowpox-based) vaccines in the past are therefore not prone to developing Ab-dependent enhancement of viral infectiousness upon subsequent exposure to MPV. However, asymptomatic human-to-human transmission of an antigenically shifted influenza variant spilling over from an animal reservoir (e.g., birds) may become particularly problematic in individuals who have previously recovered from productive infection with a common seasonal influenza virus type or who have previously been vaccinated against predominantly circulating influenza virus types (i.e., primarily the elderly and people with co-morbidities are who are otherwise immune suppressed). Zoonotic infection of these individuals with an antigenically shifted viral variant (most likely avian influenza) will likely lead to more and more cases of Ab-dependent enhancement of influenza disease[14] in humans. However, severe disease is unlikely to occur due to trained cell-based innate immunity (in C-19 unvaccinated persons) or cell-based adaptive immunity (in C-19 vaccinated persons). Should public health authorities recommend vaccination of this vulnerable group against the zoonotic influenza virus (most like, avian influenza virus), we will undoubtedly witness circulation of more infectious variants in highly vaccinated populations, resulting in enhanced rates of disease predominantly in C-19 unvaccinated children (because of a higher chance of re-infection shortly after previous exposure) and individuals who have previously been primed with common (seasonal) influenza virus types.
Similar to the epidemic predictions made for MPV, asymptomatic transmission of zoonotic influenza (most likely avian influenza) from highly C-19 vaccinated populations will likely give rise to multi-country epidemics of zoonotic influenza in poorly C-19 vaccinated populations that are immunologically naïve to the transmitted zoonotic influenza virus.
Similar also to the risks associated with MPV vaccination of young children, immunization of young children with any type of zoonotic influenza vaccine is at risk of causing (severe) zoonotic influenza disease (i.e., in the case of replication-competent vaccines) or immunopathologies (i.e., in the case of replication-incompetent vaccines).
In conclusion, no C-19 unvaccinated person should be vaccinated with common (seasonal) or zoonotic influenza virus types once human-to-human transmission of antigenically shifted (i.e., more infectious) influenza variants is occurring!
References
1. Populations aged < 50y have not been vaccinated in the past against smallpox. The smallpox vaccine uses live attenuated, replication-competent cowpox (vaccinia) virus and largely protects against monkeypox disease.
2 Infections with these viruses typically cause acute self-limiting viral disease
4 For the purpose of this manuscript, ‘vulnerable’ refers to individuals from sexual minority communities (SMCs), wherein SMCs refer to gay and bisexual male communities engaging in high-risk sexual behaviors for MPV infection (e.g., anogenital intercourses)
5 Disease in vulnerable, C-19 vaccinated individuals occurs when the virus breaks through the cytolytic immune defense provided by the hyperactivated CTLs
6 Regardless of safety concerns about potential side-effects, live attenuated, replication-competent orthopox vaccines will not be effective when used in highly C-19 vaccinated populations. This is because elimination of MPV- infected cells by cytotoxic innate or adaptive immune cells (i.e., trained innate NK cells or CTLs in the non-C19- vaccinated or C-19 vaccinated, respectively) will largely prevent ‘vaccine take’.
7 Enhanced intrinsic infectiousness could even enable airborne transmission (e.g., via particle/ droplet aerosol) as in the case of smallpox
8 Re-infection with MPV in the presence of non-neutralizing, low-affinity anti-MPV Abs enhances viral infectiousness and, therefore, disease in young, C-19 unvaccinated children
10 The additional protective effect of past vaccination with smallpox vaccines might predominantly benefit the elderly (> 65 y) and vulnerable people.
12 Herd immunity sensu stricto relates to a level of naturally induced, protective immunity that has been established in the majority of the population and is high enough to protect the remainder of that population by virtue of diminished infectious transmission.
13 Although monkey pox is an ASLVD, it can be considered an ASLVI when spreading in a highly C-19 vaccinated population at this stage of the C-19 pandemic (i.e., due to hyperactivation of cytolytic CD8+ T-cells)
14 This is because the antigenically shifted immune escape variant from the animal reservoir will not properly match the vaccine-induced Abs.
NorthShore University HealthSystem in Chicago was sued by a nonprofit religious organization called Liberty Counsel. The group claims that NorthShore violated workers’ religious autonomy by dismissing religious exemptions and forcing all workers to receive the COVID-19 vaccine. NorthShore was in the wrong and decided to settle for $10,337,500.
Other groups will follow suit. They may have granted the pharmaceutical companies immunity, but there was a grey area for employers. Countless people lost their jobs due to the vaccine mandate, which likely was a violation of the Constitution.
Lawsuits may begin with large corporations, but if the Republicans regain control, health agencies and government officials may be investigated as well. This lawsuit is a major win for medical autonomy as companies will be less likely to comply with government mandates as they now know they could risk legal retaliation.
Geert Vanden Bossche, DVM, PhD General Manager at Voice for Science and Solidarity | The biggest challenge in vaccinology: Countering immune evasion
Posted originally on TS New on Jul. 22, 2022, 9:00 a.m.
Opinion Article
Intra-pandemic vaccination of toddlers with non-replicating antibody-based vaccines targeted at ASLVI[1]– or ASLVD[2]-enabling glycosylated viruses prevents education of innate immune effector cells (NK cells).
by Geert Vanden Bossche and Rob Rennebohm
Key message:
Antibody-based vaccines teach the immune system to produce high levels of antibodies that are directed against the surface protein that is responsible for initiation of viral infection. Due to their high specificity and strong binding capacity, these vaccinal antibodies (Abs) outcompete the child’s innate antibodies for binding to the virus[3]. This not only sidelines virus-neutralization by the natural innate immune system but also hampers the ability of innate antibodies to educate the innate immune system’s NK cells (Natural Killer cells) regarding NK cell recognition of (and appropriate response to) molecular self-mimicking patterns that are expressed on virus-infected host cells. This is particularly problematic when mass vaccination campaigns are conducted during a pandemic as those drive natural selection and dominant expansion of more infectious immune escape variants.
Strong immune priming as induced by vaccines elicits long-lived Ab titers. Even in the absence of further booster shots, repeated exposure to more infectious circulating variants will recall these vaccinal antigen (Ag)-specific Abs and thereby sustain high-titer antibody responses. When immature, low-affinity Abs become exposed to the virus, (which may occur when vaccines are administered during a pandemic), these Abs may bind to the virus without neutralizing it. This in its own right could already provoke Ab-dependent enhancement of infection (ADEI) by the target virus. Vaccinated toddlers are particularly at risk of ADEI as their innate immune system has not yet been trained. Consequently, young children who are vaccinated during a pandemic with non-replicating viral vaccines (directed at ASLVI- or ASLVD-enabling glycosylated viruses[4]) are at high risk of developing severe disease.
In addition, boosting of vaccinal Abs as a result of repeated exposure to more infectious immune escape variants will lead to prolonged suspension of NK cell education in these vaccinated toddlers. When, for a prolonged period of time, NK cells are prevented from being sensitized to pathogen-derived self-mimicking peptide patterns that are expressed on infected or otherwise pathologically altered cells, they may end up becoming tolerant to these patterns, which are typically shared among several different glycosylated pathogenic agents (G. Vanden Bossche, former provisional patent application). That is, the NK cells become hyporesponsive or desensitized to these pathogenic agents[5]. This opens the door to recognition by B and T cells of traditional antigens that are naturally expressed later on in the process of infection or pathologic alteration. Recognition by these ‘foreign-centered’ effector cells may enable abrogation but not prevention of infection (i.e., in the case of infectious pathogens) or lead to immune pathology (e.g., in the case of pathologically altered autologous host cells evolving towards expression of foreign proteins).
So, when the normal NK cell responsiveness to the patterns expressed on the surface of a specific glycosylated virus-infected cell (e.g. a SARS-CoV-2 infected cell) is downregulated, so is the NK cell response to largely homologous patterns on cells infected by other glycosylated viruses. That is how – in young children- vaccinal antibody-mediated interference with the education and response of NK cells regarding one particular ASLVI- or ASLVD-enabling glycosylated virus also interferes with the education and response of their NK cells regarding other glycosylated viruses causing ASLVI or ASLVD. This renders the vaccinated young child less able to handle not only the virus it got vaccinated against but also other glycosylated viruses (of course, unless that child got previously vaccinated with childhood vaccines comprising live attenuated glycosylated viruses such as measles, mumps, rubella, varicella, rotavirus). That is why/how vaccinated young children become more susceptible to other glycosylated viruses (besides the one they got vaccinated against).
The above-mentioned prolonged suspension of NK cell education, and the associated repetitive desensitization of NK cells, will not be ‘diluted’ by a “sporadic” sensitizing event (from an intercurrent influenza infection, e.g.).
The earlier this prolonged suspension of NK cell education occurs after the maternal Abs have waned, the more detrimental the effect will be. This is because it is during early childhood, when children have their greatest and most important capacity for education and practice of their innate immune system, that they ought to exploit this opportunity to actively kick off their own immune defense against ‘foreign’ while ensuring tolerance towards ‘self’. This opportunity occurs only once in a person’s life-time! Once the functional capacity of innate Abs wanes, the instructions conveyed to ‘self-centered’ innate immune cells (i.e., NK cells) on how to recognize self-mimicking patterns associated with ‘foreign’ as opposed to ‘self’ proteins may not be strong enough to prevent irreversible priming of ‘foreign-centered’ antigen-specific B and T cells. Once such priming has occurred, the child’s immune system will have irrevocably missed that small window of opportunity to draw the thinnest possible line between ‘self’ and ‘foreign’, enabling the immune system to discriminate ‘self’ from ‘self-mimicking’ (or ‘altered self’). If one leaves it up to the adaptive immune system, a line that thin will not be drawn as the adaptive immune system has only been conceived to distinguish ‘foreign’ from ‘self’. Deficient or insufficient education of ‘self-centered’ NK cells will therefore inevitably predispose the young child to immunopathologies (ranging from allergies over inflammatory diseases up to autoreactivity).
On the other hand, once their pre-priming has been properly established, ‘training’ of NK cells can take place at any later time. Training consists of imprinting immunological memory on pre-primed NK cells following their epigenetic reprogramming. Such functional reprogramming provides these cells with sufficient plasticity to establish an “adaptive” phenotype to meet the demands and challenges of altered environmental conditions (e.g., enhanced viral infectious pressure). However, innate immune effector cells cannot be trained unless they first got educated on how to recognize potential changes/ alterations they may need to adapt to. It goes, therefore, without saying that any intervention in this delicately evolving ecosystem cannot even be considered without an in-depth understanding of the mechanisms at play and the impact thereon of the targeted immune intervention.
[3] For the purpose of this manuscript, ‘virus’ relates to an ASLVI- or ASLVD-enabling glycosylated virus
[4] Examples of glycosylated viruses [other than SARS-CoV-2] causing ASLVI or ASLVDs: seasonal influenza, RSV, dengue virus and viruses responsible for vaccine-preventable infections: measles, mumps, rubella, varicella, rotavirus or other more virulent glycosylated viruses such as zoonotic influenza (e.g., avian influenza virus), parapox virus (e.g., smallpox virus), Ebola virus, Marburg virus
[5] The biological mechanism for this fine regulation of the NK cell response is due to downregulation of germline encoded “activating receptors” on NK cells, and/or upregulation of “inhibitory receptors” on NK cells, and/or hypo-responsiveness to “activation signaling.” More details on the underlying mechanisms of this fine regulation of NK cells can be found in the literature as , for example, published by Orr, Mark T., and Lewis L. Lanier. “Natural killer cell education and tolerance.” Cell 142.6 (2010): 847-856 and Perera Molligoda Arachchige AS. Human NK cells: From development to effector functions. Innate Immunity. 2021;27(3):212-229.
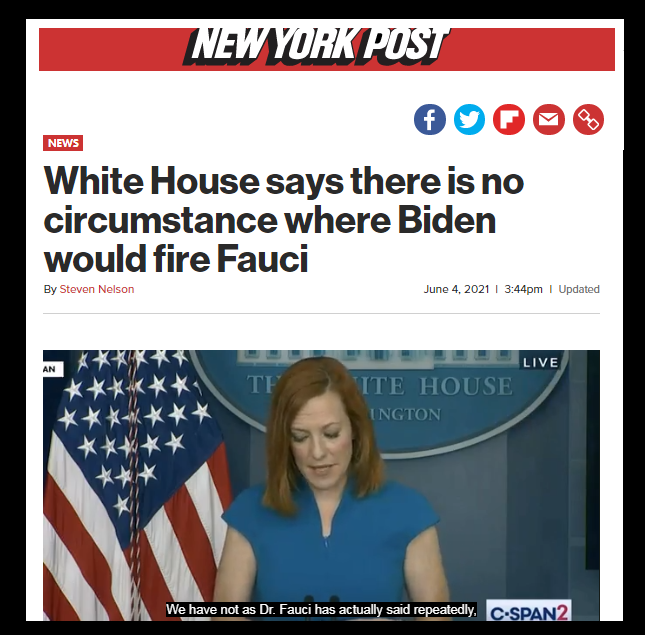
Dr. Anthony Fauci, Mr. COVID himself, announced that he would retire at the end of Joe Biden’s term. Fauci hinted in an interview that he fears an investigation into his personal dealings if the conservatives take back control next year. “They’re going to try and come after me, anyway. I mean, probably less so if I’m not in the job,” Fauci told Politico. We know the vaccination does not work and causes more harm than good. The long-term effects of the vaccines and lockdowns are now coming to light, and Fauci is responsible as the mascot for the entire COVID agenda.
Fauci maintains that boosters will be necessary every year, similar to the flu vaccine, despite the components of the two vaccines differing drastically. Fauci also admitted that the people are waking up to the fear-mongering and brainwashing techniques he used to scare the world to stay inside.
“It’s becoming more and more difficult to get people to listen, because even the people who are compliant want this behind them,” Fauci said, bewildered that people do not obey his every word. “What I try to convince them [of], with my communication method, is we’re not asking you to dramatically alter your lifestyle. We’re not asking you to really interfere with what you do with your life. We’re just asking you to consider some simple, doable mitigation methods.”
The ”simple, doable mitigation methods” involved involuntary house arrest, school closures, business closures, and forced vaccinations of an EXPERIMENTAL mRNA gene-altering therapy. These policies were a gut punch to the global economy. Countless people lost their jobs and lives due to his warped God-complex view of “trust the science.” This man should be held accountable for the damage he has done to society – permanent damage that will linger for generations to come.
I have created this site to help people have fun in the kitchen. I write about enjoying life both in and out of my kitchen. Life is short! Make the most of it and enjoy!
This is a library of News Events not reported by the Main Stream Media documenting & connecting the dots on How the Obama Marxist Liberal agenda is destroying America